Анатомия растений
Анатомия растений, раздел ботаники, посвященный изучению внешнего и внутреннего
строения растений. Основной объект этой науки - т.н. сосудистые растения,
обладающие специализированной водопроводящей тканью - ксилемой. К ним относятся
плауны, хвощи, папоротники, голосеменные и покрытосеменные (цветковые) растения.
Исторический аспект. Первые анатомические описания растений встречаются в
трудах Теофраста (3 в. до н.э.). Он различал такие структурные части, как
корень, стебель, ветвь, цветок и плод, а главными растительными тканями считал
кору, древесину и сердцевину. Эти представления в общих чертах сохранились до
сегодняшнего дня. Когда Р.Гук в 1665 с помощью сконструированного им микроскопа
открыл клетку, это создало новые возможности для изучения анатомии растений. Н.
Грю в 1682 в своей прекрасно иллюстрированной работе описал микроскопическое
строение различных растительных структур, усмотрев в нем сходство с
переплетением нитей в ткани. Х.фон Моль в 1831 проследил ход проводящих пучков
в стеблях, корнях и листьях. К.Санио в 1863 выяснил происхождение камбия и
показал, каким образом ежегодно образуется новый цилиндр ксилемы и флоэмы
(ткани, по которой в растении транспортируются органические вещества). В 1877
Антон де Бари опубликовал свой классический труд Сравнительная анатомия
вегетативных органов явнобрачных и папоротников (Vergleichende Anatomie der
Vegetationsorgane der Phanerogamen und Farne), в котором суммировал весь
накопившийся к тому времени материал по этому вопросу. В 20 в. развитие
анатомии растений шло особенно бурными темпами и было неразрывно связано с
общим прогрессом биологических наук, обусловленным появлением новых методов
исследования.
При чрезвычайном внешнем разнообразии растений их клетки имеют сходное
устройство. Чтобы понять внутреннее строение организма, необходимо
познакомиться с общей организацией его клеток и с особенностями различных их
типов.
Клетка. Растительная клетка состоит из студенистой протоплазмы, окруженной
жесткой оболочкой (клеточной стенкой), а последняя - главным образом из
секретируемых протоплазмой целлюлозы и пектиновых веществ. Во многих клетках
после завершения их роста на внутренней стороне исходной (первичной) клеточной
стенки откладывается т.н. вторичная. Протоплазма - это смесь воды, белков,
сахаров, жиров, кислот, солей и многих других веществ. Распределенные в
правильных соотношениях по различным частям клетки, они обеспечивают протекание
биохимических процессов, т.е. жизненных функций. Под микроскопом видно, что
протоплазма подразделяется на ядро и цитоплазму, в которой находятся пластиды.
Ядро - это более или менее сферическое тельце, окруженное двойной мембраной.
Оно координирует химические процессы в клетке и содержит ее наследственный
материал. Цитоплазма - вязкое вещество, содержащее сложную сеть структур и
более крупные образования, в т.ч. свойственные только растениям пластиды. В
бесцветных пластидах (лейкопластах) запасаются питательные вещества, в зеленых
(хлоропластах) идет фотосинтез сахаров. В старых клетках центральную часть
занимает вакуоль - окруженное мембраной скопление водянистой жидкости, в
которой растворены различные вещества. При этом протоплазма оттеснена на
периферию в виде тонкого слоя, примыкающего к клеточной стенке. От клеток с
описанным выше строением ведут свое происхождение все прочие их типы,
встречающиеся в растениях.
Ткани. Растительный организм подразделяется на специализированные зоны, или
части, особенности которых определяются типами и взаиморасположением клеток,
входящих в их состав. Такие участки называются тканями. Согласно классическому
определению, ткани различаются происхождением, структурой и функциями. Однако
они не всегда четко отграничены друг от друга, не обязательно однородны, а их
функции могут частично совпадать. Это чрезвычайно затрудняет классификацию
тканей, поэтому в наше время все чаще говорят просто о различных зонах растения.
В таком топографическом смысле на поперечном срезе стебля и корня сосудистого
растения от периферии к центру обычно можно различить следующие концентрические
зоны: эпидермис (эпидерму), кору, проводящий цилиндр и (часто) центральную
сердцевину.
Корень - это безлистная часть растения, поглощающая из почвы (или другой среды)
воду с растворенными в ней питательными веществами, удерживающая его в
субстрате, а иногда служащая главным запасающим органом, например у моркови или
свеклы. Топографически в нем четко видны эпидермис, кора и стела (осевой, или
центральный, цилиндр); они растут в результате деления и дифференцировки клеток
апикальной (верхушечной) меристемы. Меристемами называют группы клеток,
сохраняющие способность к делению и образованию специализированных клеток (уже
не делящихся). Апикальная меристема отделяет с поверхности корневой чехлик,
предохраняющий ее от механических повреждений в ходе продвижения кончика корня
сквозь почву. Деление, рост и дифференцировка клеток - это последовательные
процессы, которые позволяют различать по вертикали следующие зоны: корневой
чехлик, апикальную меристему, зону растяжения и зону созревания. В них
прослеживаются различные стадии формирования эпидермиса, коры и стелы.
Непосредственно над зоной растяжения эпидермальные клетки образуют длинные
цилиндрические выросты - корневые волоски. Они увеличивают всасывающую
поверхность корня.
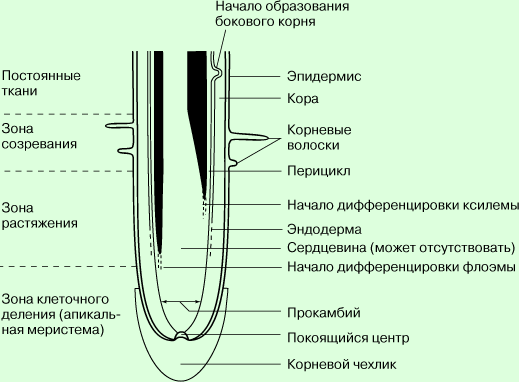
Структура типичного корня в участке роста (продольный разрез). Рост корня
происходит к результате деления, роста и дифференцировки клеток <образующей> ткани -
апикальной (верхушечной) меристемы. Эта ткань составляет зону клеточного
деления. Однако и сама она представлена несколькими зонами, клетки которых
различаются по своей дальнейшей судьбе. Покоящийся центр - группа
меристематических клеток-предшественников, делящихся медленнее, чем все другие,
образованные ими клетки апикальной меристемы. Прокамбий - зона апикальной
меристемы, дающей начало центральному цилиндру. Последний включает проводящие
ткани - ксилему и флоэму, - сердцевину (иногда), а также перицикл (слой
меристемы, образующий боковые корни) и камбий (вторичную меристему,
обеспечивающую рост корня в толщину). Апикальная меристема, окружающая
прокамбий, состоит из двух зон, одна из которых образует эпидермис, а другая -
кору с эндодермой. Зона растяжения (роста) клеток на рисунке укорочена. Заметим,
что она частично перекрывается зоной созревания (дифференцировки) клеток.
Стела состоит из первичной ксилемы и первичной флоэмы, начало которым дает
апикальная меристема. Ксилема находится ближе к центру и образует радиально
уплощенные лучи, которые чередуются с тяжами флоэмы. Сердцевина в корнях по
большей части отсутствует, но у однодольных встречается чаще, чем у двудольных.
Боковые корни закладываются в слое меристематических клеток (перицикле) на
поверхности стелы и пробивают себе путь наружу через кору. В корнях, способных
расти в толщину (вторично утолщаться), между ксилемой и флоэмой развивается
слой вторичной меристемы - камбия. Деление и дифференцировка его клеток дают
вторичную ксилему (с внутренней стороны) и вторичную флоэму (с наружной). При
усиленном росте в толщину эпидермис и кора разрываются и отмирают. К этому
моменту во внутренней части первичной коры, перецикле или вторичной флоэме
закладывается пробковый камбий (феллоген), образующий вокруг корня защитный
слой пробки.
Стебель - это осевая часть растения, которая несет листья и репродуктивные
органы. Он служит опорой надземным органам, обеспечивает поступление в них воды,
а также транспорт синтезированных питательных веществ к корням и в другие места,
где используются эти соединения. Стебли могут быть зелеными, как у кактусов, т.
е. способными к фотосинтезу. Важную роль они играют и в запасании питательных
веществ, а нередко служат для вегетативного размножения, например у сахарного
тростника или картофеля.
Апикальная меристема. Верхушка стебля прикрыта, как колпачком, массой делящихся
недифференцированных клеток - апикальной меристемой, образующей конус
нарастания. Здесь возникают зачатки листьев, которые сначала черепицеобразно
налегают друг на друга. Постепенно промежутки между последовательно
появляющимися листьями, растягиваясь, превращаются в междоузлия, а те места,
где листья отходят от зрелого стебля, становятся узлами.
Стела изучена лучше, чем любая другая часть растения. Различают два основных ее
типа. Протостела состоит из сплошного тяжа ксилемы, окруженного флоэмой;
сифоностела отличается тем, что ксилема образует полый цилиндр, окружающий
центральную сердцевину. С эволюционной точки зрения, протостела примитивнее.
Она характерна для стеблей и корней плаунов, некоторых папоротников, первых
наземных растений (судя по ископаемым остаткам) и корней семенных видов. Во
всех остальных случаях встречается сифоностела (с теми или иными модификациями).
От стелы в листья идут сосудистые пучки, называемые листовыми следами. Над
листовым следом первичные ксилема и флоэма не развиваются. Здесь в центральном
цилиндре находятся т.н. листовые прорывы (листовые щели), заполненные
паренхимой. В эволюции сосудистых растений прослеживается тенденция к
увеличению высоты листовых прорывов, в результате чего стела приобретает вид
ажурного цилиндра из отдельных сосудистых пучков. Это т.н. диктиостела. Она
часто встречается у травянистых семенных растений. Дальнейшая специализация
прослеживается у однодольных: сосудистые пучки столь многочисленны, что уже не
располагаются цилиндром вокруг сердцевины, а рассеяны по всей толще стебля.
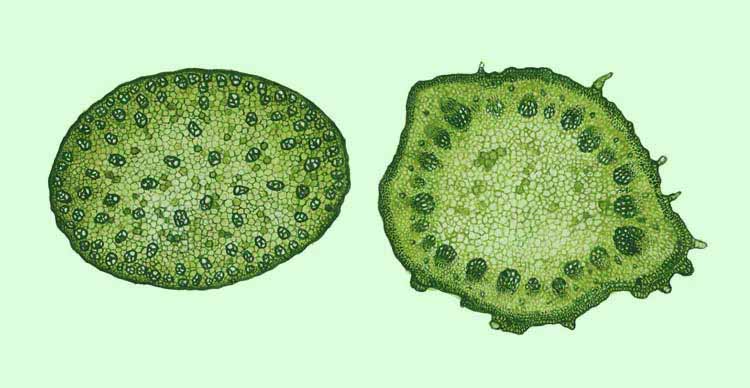
Сосудистые пучки у однодольных рассеяны по всей толще стебля (слева); у
двудольных они обычно расположены кольцом, окружающим сердцевину (справа).
У всех семенных растений, за исключением однодольных и некоторых
высокоспециализированных форм, между первичными ксилемой и флоэмой формируется
камбий. У древесных пород умеренной климатической зоны ежегодно в течение
вегетационного периода образуется хорошо заметное годичное кольцо ксилемы,
состоящее из двух слоев древесины - ранней (весенней) и поздней (летней).
Лист по своему внутреннему строению совершенно не похож ни на стебель, ни на
корень. Сверху он покрыт слоем эпидермиса. Под ним находится т.н. палисадный
мезофилл из одного или нескольких слоев клеток, вытянутых перпендикулярно
плоскости листа. Далее следует многослойный губчатый мезофилл, клетки которого
образуют трехмерную сеть с хорошо развитыми межклетниками. Нижнюю поверхность
листа покрывает эпидермис, пронизанный устьицами. Палисадный и губчатый
мезофилл - это по существу видоизмененная кора, а листовые жилки - ответвления
стелы. Листья же можно считать уплощенными и укороченными ветвями. См. также
Лист.
Цикл углерода
Цикл углерода, круговорот углерода, - циклическое перемещение углерода между
миром живых существ и неорганическим миром атмосферы, морей, пресных вод, почвы
и скал. Это один из важнейших биогеохимических циклов, включающий множество
сложных реакций, в ходе которых углерод переходит из воздуха и водной среды в
ткани растений и животных, а затем возвращается в атмосферу, воду и почву,
становясь снова доступным для использования организмами. Поскольку углерод
необходим для поддержания любой формы жизни, всякое вмешательство в круговорот
этого элемента влияет на количество и разнообразие живых организмов, способных
существовать на Земле.
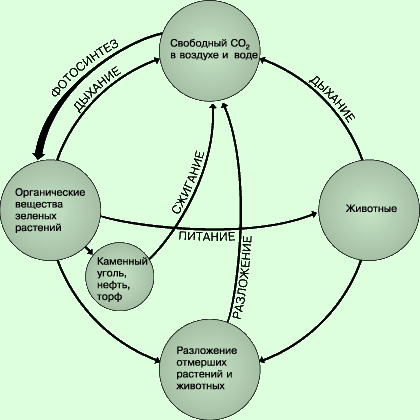
Циклом углерода называют круговорот этого элемента между живыми организмами и
неорганической средой. На схеме показаны основные процессы цикла.
Источники и резервы углерода. Основной источник углерода для живых организмов -
это атмосфера Земли, где данный элемент присутствует в виде диоксида углерода
(углекислого газа, СО2). В течение многих миллионов лет концентрация СО2 в
атмосфере, по-видимому, существенно не менялась, составляя ок. 0,03% веса
сухого воздуха на уровне моря. Хотя доля СО2 невелика, его абсолютное
количество поистине огромно - ок. 750 млрд. т. В атмосфере СО2 переносится
ветрами как в вертикальном, так и в горизонтальном направлениях.
Диоксид углерода присутствует в воде, где он легко растворяется, образуя слабую
угольную кислоту Н2СО3. Эта кислота вступает в реакции с кальцием и другими
элементами, образуя минералы, называемые карбонатами. Карбонатные породы,
например известняк, находятся в равновесии с диоксидом углерода, который
содержится в контактирующей с ними воде. Аналогичным образом количество СО2,
растворенного в океанах и пресных водах, определяется его концентрацией в
атмосфере. Общее количество растворенных и осадочных углеродсодержащих веществ
оценивается примерно в 1,8 трлн.т.
Углерод в соединении с водородом и другими элементами является одним из
основных компонентов клеток растений и животных. Например, в организме человека
он составляет ок. 18% массы тела. Многочисленность и очень широкое
распространение живых организмов не позволяют удовлетворительно оценить общее
содержание в них углерода. Можно, однако, приблизительно оценить суммарное
количество углерода, связываемого растениями, а также выделяемого в процессе
дыхания растений, животных и микроорганизмов. Установлено, что зеленые растения
поглощают в год ок. 220 млрд.т CO2. Почти такое же количество этого вещества
выделяется в неорганическую среду в процессе дыхания всех живых организмов, а
также в результате разложения и сгорания органических веществ.
При определенных условиях разложения и сгорания созданных живыми организмами
веществ не происходит, что ведет к накоплению углеродсодержащих соединений. Так,
например, древесина живых деревьев может быть на 3-4 тысячелетия надежно
защищена от микробного разложения и от пожара корой, способной противостоять
действию микробов и огня. Древесина же, попавшая в торфяное болото, сохраняется
еще дольше. В обоих случаях связанный в ней углерод оказывается как бы в
ловушке и надолго выводится из круговорота. В условиях, когда органическое
вещество оказывается захороненным и изолированным от воздействия воздуха, оно
разлагается только частично и содержащийся в нем углерод сохраняется. Если
впоследствии в течение миллионов лет эти органические остатки подвергаются
давлению вышележащих отложений и нагреванию за счет земного тепла, значительная
часть его превращается в ископаемое топливо, например в каменный уголь или
нефть. Ископаемое топливо образует природный резерв углерода. Несмотря на
интенсивное его сжигание, начавшееся с 1700-х годов, неизрасходованными еще
остаются примерно 4,5 трлн.т.
Фотосинтез. Основной путь, посредством которого углерод из мира неорганического
перемещается в мир живого, - это осуществляемый зелеными растениями фотосинтез.
Данный процесс представляет собой цепь реакций, в ходе которых растения
поглощают из атмосферы или воды диоксид углерода, связывая его молекулы с
молекулами специального вещества - акцептора СО2. В ходе других реакций, идущих
с потреблением солнечной (световой) энергии, происходит расщепление молекул
воды и использование высвобождающихся ионов водорода и связанного СО2 в синтезе
богатых углеродом органических веществ, в том числе акцептора СО2.
На каждую молекулу СО2, которую поглощает растение, чтобы синтезировать
органические вещества, выделяется молекула кислорода, образованная при
расщеплении воды. Предполагается, что именно таким путем образовался весь
свободный кислород атмосферы. Если бы процесс фотосинтеза на Земле внезапно
прекратился и нарушился углеродный цикл, то, согласно имеющимся расчетам, весь
свободный кислород исчез бы из атмосферы примерно за 2000 лет. См. также
Фотосинтез.
Другие реакции. Зеленое растение использует углерод образуемых им органических
веществ разными способами. Например, он может накапливаться в составе крахмала,
запасаемого в клетках, или целлюлозы - основного структурного материала
растений и питательного вещества для многих других организмов. И крахмал и
целлюлоза усваиваются в качестве пищи только после расщепления на составляющие
их 6-углеродные сахара (т.е. сахара, содержащие по шесть атомов углерода в
молекуле). В отличие от крахмала - нерастворимого высокомолекулярного
соединения - 6-углеродные сахара легко растворимы и, перемещаясь по растению,
служат источником энергии и материалом для роста и обновления клеток, а также
для их восстановления в случае повреждений. Проростки, например, расщепляют
запасенные в семени крахмал и жиры, получая из них более простые органические
вещества, используемые в процессе клеточного дыхания (для высвобождения их
энергии) и для роста.
У животных поглощенная пища подвергается аналогичному процессу переваривания.
Прежде чем ее основные компоненты могут быть усвоены, они должны быть
преобразованы: углеводы - в 6-углеродные сахара, жиры - в глицерин и жирные
кислоты, белки - в аминокислоты. Эти продукты переваривания служат животному
источниками энергии, высвобождаемой при дыхании, а также строительными блоками,
необходимыми для роста организма и обновления его компонентов. Подобно
растениям, животные способны переводить питательные вещества в форму, удобную
для запасания. Аналог крахмала у животных - это гликоген, образуемый из
излишков 6-углеродных сахаров и накапливаемый в качестве энергетического
резерва в печени и мышечных клетках. Избыток сахара может превращаться также в
жирные кислоты и глицерин, которые вместе с такими же веществами, поступающими
с пищей, используются для синтеза жиров, накапливаемых в ткани. Таким образом,
процессы синтеза обеспечивают запасание богатых углеродом и связанной энергией
веществ, что позволяет организму выживать в периоды нехватки пищи.
Одна из характерных особенностей всего живого - постоянная потребность в
энергии. Организм получает энергию посредством дыхания - целой серии процессов,
в ходе которых сложные углеродсодержащие молекулы превращаются в простые.
Большинство растений и животных способно только к аэробному дыханию, т.е. они
поглощают кислород из воздуха, образуя диоксид углерода и воду в качестве
конечных продуктов. Однако существуют некоторые бактерии, простейшие и даже
многоклеточные животные (кишечные паразиты), являющиеся анаэробами: они
способны жить в отсутствие кислорода в среде; при этом конечными продуктами их
анаэробного дыхания (брожения разных типов) тоже служит диоксид углерода и вода.
Очень немногие организмы (например, дрожжи) могут быть как аэробами, так и
анаэробами. В аэробных условиях дрожжи образуют в качестве конечных продуктов
диоксид углерода и воду, а в анаэробных - диоксид углерода и этиловый спирт.
Таким образом, независимо от типа дыхания оно всегда ведет к высвобождению
углерода в форме диоксида, который затем снова вовлекается в глобальный цикл.
После своей смерти растения и животные становятся пищей для т.н. редуцентов -
организмов, осуществляющих разложение органического вещества. Большая часть
редуцентов представлена бактериями и грибами, клетки которых выделяют наружу, в
свое непосредственное окружение, небольшие количества пищеварительной жидкости,
расщепляющей субстрат, а затем потребляют продукты такого "переваривания". Как
правило, редуценты имеют ограниченный набор ферментов и соответственно
используют в качестве пищи и источника энергии только немногие типы
органических веществ. Обычные дрожжи, например, перерабатывают только 6- и 12-
углеродные сахара, содержащиеся в разрушенных клетках перезрелых фруктов или в
густом (с мякотью) соке, полученном при их раздавливании. Однако при
достаточной длительности воздействия разнообразных редуцентов все
углеродсодержащие вещества растений или животных в конце концов разрушаются до
диоксида углерода и воды, а высвобожденная энергия используется организмами,
осуществляющими разложение. Многие искусственно синтезированные органические
соединения тоже подвержены биологическому разрушению (биодеградации) - процессу,
в ходе которого редуценты получают энергию и необходимый строительный материал,
а в атмосферу выделяется углерод в форме диоксида углерода.
Фотосинтез
Елена Савинкина
Фотосинтез - образование органических веществ зелеными растениями и некоторыми
бактериями с использованием энергии солнечного света.
В ходе фотосинтеза происходит поглощение из атмосферы диоксида углерода и
выделение кислорода.
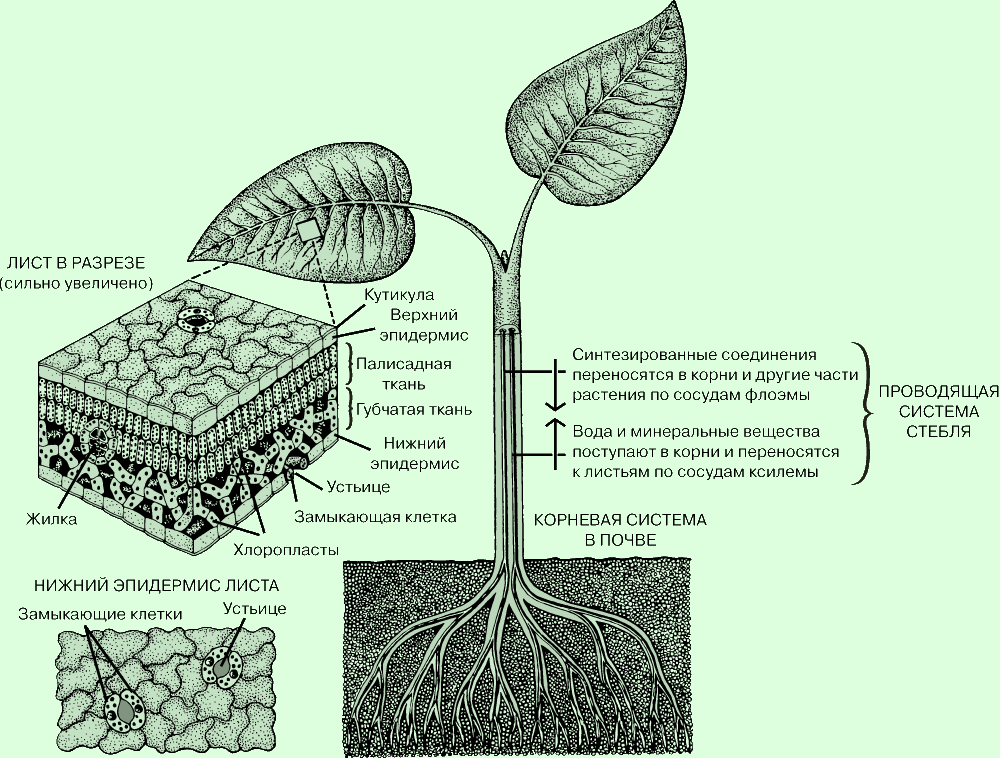
Фотосинтез - это процесс, от которого зависит вся жизнь на Земле. Он происходит
только в растениях. В ходе фотосинтеза растение вырабатывает из неорганических
веществ необходимые для всего живого органические вещества. Диоксид углерода,
содержащийся в воздухе, проникает в лист через особые отверстия в эпидермисе
листа, которые называют устьицами; вода и минеральные вещества поступают из
почвы в корни и отсюда транспортируются к листьям по проводящей системе
растения. Энергию, необходимую для синтеза органических веществ из
неорганических, поставляет Солнце; эта энергия поглощается пигментами растений,
главным образом хлорофиллом. В клетке синтез органических веществ протекает в
хлоропластах, которые содержат хлорофилл. Свободный кислород, также
образующийся в процессе фотосинтеза, выделяется в атмосферу.
Первым обнаружил, что растения выделяют кислород, английский химик и философ
Джозеф Пристли около 1770. Вскоре было установлено, что для этого необходим
свет и что кислород выделяют только зеленые части растений. Затем исследователи
нашли, что для питания растений требуется диоксид углерода и вода, из которых
создается большая часть массы растений. В 1817 французские химики Пьер Жозеф
Пелатье (1788-1842) и Жозеф Бьенеме Каванту (1795-1877) выделили зеленый
пигмент хлорофилл (по-гречески croz - chloros, зеленый; julln - phyllon, лист).
Позднее российский ученый Климент Аркадьевич Тимирязев (1843-1920) показал, что
фотосинтез проходит с наибольшей интенсивностью в тех областях солнечного
спектра, где находятся максимумы поглощения хлорофилла.
К середине 19 в. было установлено, что фотосинтез является процессом, как бы
обратным дыхательному. Французский ученый Жан Батист Буссенго (1802-1887) в
своих работах, опубликованных в это время, утверждал, что в процессе
фотосинтеза происходит выделение кислорода из углекислого газа. Это мнение в
научной литературе господствовало длительное время.
В 1860-х было высказано предположение, что диоксид углерода в растениях
восстанавливается до органических кислот, в частности, муравьиной и щавелевой.
Затем эти кислоты при дальнейшем восстановлении переходят в углеводы. В 1861
русский химик Александр Михайлович Бутлеров получил при действии известковой
воды на формальдегид сиропообразное вещество, содержащее углеводы. Основываясь
на этом открытии, немецкий химик Адольф Байер в 1870 высказал предположение,
что первичным продуктом восстановления диоксида углерода в зеленых растениях
является формальдегид, который затем превращается в углеводы. Эта гипотеза
привлекла всеобщее внимание - она казалась наиболее правдоподобной. Однако она
ничего не говорила о механизме выделения кислорода.
Этим вопросом занялся в конце 19 в. биохимик Алексей Николаевич Бах (1857-1946).
На основе экспериментальных исследований он пришел к выводу, что при
ассимиляции диоксида углерода источником выделяющегося молекулярного кислорода
являются пероксиды, образующиеся из воды. Он же высказал предположение о
биокаталитической роли белков-ферментов в фотосинтезе.
В 20 в. было установлено, что процесс фотосинтеза начинается на свету в
фоторецепторах хлорофиллов, однако многие из последующих стадий могут протекать
в темноте. Общий процесс является эндотермическим (DH° ~ 469 кДж/моль СО2). В
нем участвует несколько типов хлорофилла, а также другие комплексы магния,
железа и меди.
В 1941 американский биохимик Мелвин Калвин (1911-1997) показал, что первичный
процесс фотосинтеза заключается в фотолизе молекул воды, в результате чего
образуются кислород, выделяющийся в атмосферу, и водород, идущий на
восстановление диоксида углерода до органических веществ. Используя
радиоактивный изотоп углерода 14С, бумажную хроматографию и классические методы
органической химии, Калвин и его группа смогли проследить биосинтетические пути
фотохимических процессов. К 1956 стал ясным полный путь превращения углерода
при фотосинтезе. За исследования в области ассимиляции диоксида углерода в
растениях Калвин был удостоен в 1961 Нобелевской премии по химии.
Полная последовательность всех стадий фотосинтеза пока еще выяснена не до конца,
однако интенсивная научная работа в этом направлении продолжается. Исследуется
механизм электронного транспорта, продолжается выяснение природы комплекса,
катализирующего образование кислорода, изучается структура реакционных центров
и светособирающих комплексов.
В целом, химический баланс фотосинтеза может быть представлен в виде простого
уравнения:
6CO2 + 6H2O = C6H12O6 + 6O2
Водород, необходимый для восстановления диоксида углерода до глюкозы, берется
из воды, а выделяющийся в ходе фотосинтеза кислород является побочным продуктом.
Процесс нуждается в энергии света, так как вода сама по себе не способна
восстанавливать диоксид углерода.
В светозависимой части фотосинтеза (световой реакции) происходит расщепление
молекул воды с образованием протонов, электронов и атома кислорода. Электроны,
возбужденные энергией света, восстанавливают никотинадениндинуклеотидфосфат
(НАДФ). Образующийся НАДФ-Н является подходящим восстановителем для перевода
диоксида углерода в органические соединения. Кроме того, в световой реакции
образуется аденозинтрифосфат (АТФ), который также необходим для фиксации
диоксида углерода.
В световых реакциях электроны переносятся по электрон-транспортной цепи от
одной окислительно-восстановительной системы к другой. Возбуждение электронов
для восстановления никотинадениндинуклеотидфосфата - сложный фотохимический
процесс. Он происходит в реакционных центрах (фотосистемах), которые
представляют собой белковые комплексы, содержащие множество молекул хлорофилла
и других пигментов. Только около 1% молекул хлорофилла участвуют
непосредственно в фотохимическом переносе электронов. Основная часть связана с
другими пигментами в так называемом комплексе светособирающей антенны. Энергия
кванта света, накопленного в комплексе, передается на реакционный центр, где и
используется.
Последующие процессы могут протекать в темноте (темновая реакция). Полная
последовательность превращения диоксида углерода в органические соединения
называется циклом Калвина.
В зеленых водорослях и высших растениях фотосинтез происходит в хлоропластах,
которые окружены двумя мембранами и содержат собственную ДНК. Световые реакции
катализируются ферментами, находящиеся в сложенных стопками утолщенных
мембранных мешках, а темновые реакции происходят во внутреннем пространстве
хлоропластов.
Таким образом, в основе фотосинтеза лежит превращение электромагнитной энергии
света в химическую энергию. Эта энергия, в конце концов, дает возможность
превращать диоксид углерода в углеводы и другие органические соединения с
выделением кислорода.
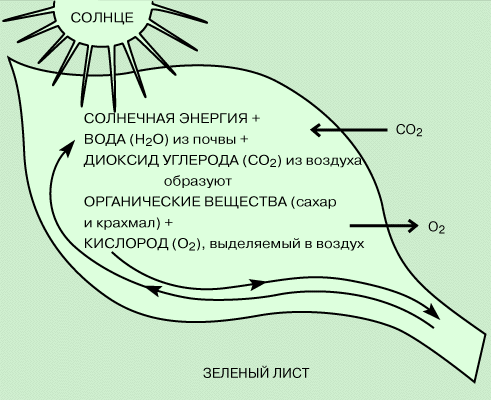
Схема фотосинтеза.
Фотосинтез, являющийся одним из самых распространенных процессов на Земле,
обуславливает природные круговороты углерода, кислорода и других элементов и
обеспечивает материальную и энергетическую основу жизни на нашей планете.
Фотосинтез является единственным источником атмосферного кислорода.
Процесс фотосинтеза является основой питания всех живых существ, а также
снабжает человечество топливом (древесина, уголь, нефть), волокнами (целлюлоза)
и бесчисленными полезными химическими соединениями. Из диоксида углерода и воды,
связанных из воздуха в ходе фотосинтеза, образуется около 90-95% сухого веса
урожая. Остальные 5-10% приходятся на минеральные соли и азот, полученные из
почвы.
Человек использует около 7% продуктов фотосинтеза в пищу, в качестве корма для
животных и в виде топлива и строительных материалов.
Лист
Лист, орган растения, исходно специализированный для осуществления фотосинтеза,
т.е. питания организма, но в ходе эволюции иногда утрачивающий эту функцию или
приобретающий дополнительные функции. Из всех творений природы зеленый, т.е.
содержащий хлорофилл, лист - наиболее важная для жизни на Земле структура. Без
нее не могли бы существовать человек и другие организмы. Атмосферный запас
кислорода пополняется непрерывным выделением этого газа из листа зеленых
растений. В год листья поглощают до 400 млрд. т углекислого газа, связывая при
этом в составе органических соединений 100 млрд. т углерода. Именно эти
органические соединения, образующиеся в листьях, служат первичным источником
пищи и жизненно важных витаминов для человека и всех диких и домашних животных.
См. также Цикл углерода.
Листья обеспечивают людей не только кислородом и пищей. В тропиках, например,
до сих пор живут в хижинах, крытых листьями пальм. Во всем мире одним из
важнейших строительных материалов остается древесина, которая не могла бы
образоваться, не будь на деревьях листьев. Если отвлечься от чисто утилитарных
нужд, следует вспомнить и о том, что листья делают нашу жизнь более приятной и
комфортабельной. Из них готовят вкусные и тонизирующие напитки, например
обычный чай из листьев чайного куста или "матэ" из листьев падуба парагвайского
- кустарника, растущего по берегам рек в Аргентине, Парагвае и на юге Бразилии.
Курение листьев табака (Nicotiana tabacum) многим помогает расслабиться. Из
листьев различных растений, например коки, наперстянки, красавки, получают
сильнодействующие лекарства. Листья алоэ настоящего (Aloe vera) содержат
вещества, излечивающие некоторые дерматиты, облегчающие боль при радиационных и
солнечных ожогах и смягчающие кожу. Некоторые листья, обладающие приятным
ароматом, используются непосредственно как приправы или служат сырьем для
производства душистых экстрактов. Именно такое применение находят, например,
листья базилика, лавра, майорана, тимьяна, лаванды и перечной мяты. Из листьев
сансевьеры цилиндрической (Sansevieria cylindrica) и агавы сизалевой (Agave
sisalana) получают волокно для изготовления канатов, из листьев некоторых
других видов плетут циновки, покрывала и шляпы.
Основные части и общие характеристики. Типичный лист состоит из трех частей:
пластинки, черешка и прилистников - мелких листовидных структур у основания
черешка. Основная часть - пластинка, обычно тонкая, плоская и зеленая. Однако у
некоторых растений ее окраска иная, например темно-красная у популярной среди
цветоводов ирезины Хербста (Iresine herbstii), пестрая у колеуса (крапивки),
или серебристая у сантолины кипарисовниковидной (Santolina chamaecyparissus),
известной также как кипарисная трава. Иногда поверхность листа опушена, т.е.
покрыта волосками - выростами наружных клеток.
Черешки некоторых листьев, например сельдерея и ревеня, очень крупные и
употребляются в пищу. Иногда же черешки вообще отсутствуют, и пластинка листа
прикреплена непосредственно к стеблю. Такие листья называют сидячими. Они
свойственны, в частности, диервилле сидячелистной (Diervilla sessilifolia),
относящейся к семейству жимолостных. Прилистники у большинства растений мелкие,
однако иногда они вполне сопоставимы по размерам с листовой пластинкой, как у
гороха огородного или хеномелеса японского. В некоторых случаях, например у
белой акации (Robinia pseudoacacia), прилистники преобразованы в шипы.
Форма листа - один из отличительных признаков вида растения. Лист бывает
простым или сложным, т.е. состоящим из нескольких листочков, в зависимости от
того, одна у него пластинка или несколько. Так, у берез, буков, ильмов, дубов и
платанов листья простые, а у конских каштанов, белой акации, шиповника, айланта
и грецкого ореха - сложные. Сложные листья бывают перисто- и пальчатосложными.
В первом случае листочки расположены двумя супротивными рядами вдоль общей оси,
как, например, у белой акации и грецкого ореха, а во втором - отходят от одной
точки, как, скажем, у конского каштана или клевера.
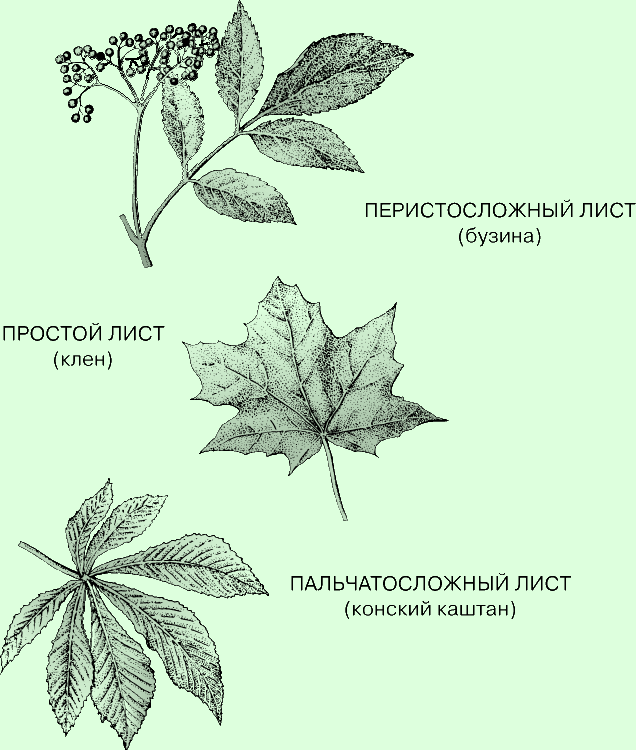
Простые и сложные листья. Лист называют простым или сложным в зависимости от
того, одна у него пластинка или несколько. Во втором случае лист может быть
перистосложным, если составляющие его листочки расположены в два ряда на общей
оси, или пальчатосложными, когда они выходят из одной точки - верхушки черешка.
Размеры листьев широко варьируют в зависимости от таксона и даже в пределах
одного вида растений. Они могут достигать в длину 20 м, например у пальмы
Raphia ruffia, произрастающей в тропической Африке и на Мадагаскаре. Очень
мелкие листья у спаржи овощной (Asparagus officinalis var. altilis), казуарины
хвощевидной (Casuarina equisetifolia) и тамариска, или гребенщика (Tamarix spp.
).
В большинстве случаев листья широкие и плоские, однако иногда они
цилиндрические, как у лука, игловидные, как у сосен, или чешуевидные, как у
кипарисов. Бывают листья линейные (у злаков), округлые (у настурции),
яйцевидные (у каркаса), сердцевидные (у липы), ланцетные (у ивы) и т.п. Иногда
наблюдается т.н. гетерофилия ("разнолистность") - на одном и том же растении
образуются разные по форме листья; например, у сассафраса лекарственного их
насчитывают пять вариантов.
Листья с ровными краями называют цельнокрайними. Среди деревьев такие листья
можно видеть, например, у кизила, сирени, рододендрона, эвкалипта, дубов
черепитчатого, иволистного и виргинского. Во многих случаях края листовой
пластинки лопастные, рассеченные, зубчатые, выемчатые. Например, у дуба
красного листья перистолопастные с шиповидными выступами жилок на вершинах
долей, а у дуба белого листья перистолопастные или плавновыемчатые без острых
углов.
У большинства растений листорасположение очередное, или спиральное: листья, как
и почки с боковыми побегами, отходят по одному от каждого узла то с одной, то с
другой стороны стебля. Пример - все березы, ильмы, дубы и орехи. У некоторых
видов, в частности кленов, калины и кизила, листья, почки и боковые побеги
расположены супротивно - с противоположных сторон каждого узла. Когда от узла
отходит три листа или более, листорасположение называют мутовчатым. В любом
случае листья отходят от стебля так, чтобы минимально затенять друг друга. Они
образуют в пространстве своего рода "листовую мозаику", призванную улавливать
как можно больше падающего на растение солнечного света.
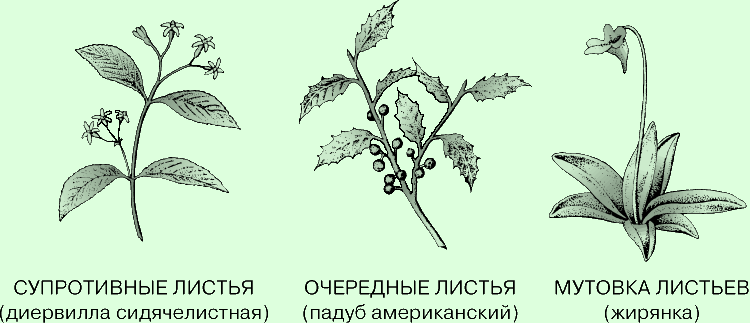
Типы листорасположения. Основных типов листорасположения - три: супротивное,
очередное (спиральное) и мутовчатое. В первом случае от каждого узла с двух
противоположных сторон стебля отходит по листу. Во втором случае листья отходят
от узлов по одному - то с одной, то с другой стороны стебля. Если от узла
отходит три листа и более, их расположение называют мутовчатым.
Водные растения
Г.И.Поплавская
Водные растения, растения, произрастающие в воде. Среди них различают гидрофиты
- растения, погружённые в воду только нижней частью, и гидатофиты - растения,
полностью или большей своей частью погруженные в воду. Обитание в водной среде
обусловило особые черты организации водных растений: значительное увеличение
поверхности тела в сравнении с его массой, что облегчает поглощение необходимых
количеств кислорода и других газов, которых в воде содержится меньше, чем в
воздухе. Увеличение поверхности тела достигается развитием больших тонких
листьев (некоторые рдесты), расчленением листовой пластинки на тонкие
нитевидные участки (уруть, роголистники, водные лютики); продырявливанием
листьев или сильным развитием воздухоносных полостей и больших межклетников. У
водных растений сильно развита разнолистность (гетерофиллия): подводные,
плавающие и воздушные листья на одном и том же растении значительно различаются
как по внутреннему, так и по внешнему строению. Так, подводные листья не имеют
устьиц; у плавающих на поверхности воды листьев устьица находятся только на
верхней стороне, у воздушных листьев устьица - на обеих сторонах. Большая
плотность водной среды обусловливает слабое развитие механических элементов в
листьях и стеблях водных растений; немногочисленные механические элементы,
имеющиеся в стеблях, расположены ближе к центру, что придаёт им большую
гибкость. Так как интенсивность света в воде резко снижается, у многих водных
растений в клетках эпидермиса имеются хлорофилловые зёрна. У водные растений
слабо развиты или даже отсутствуют сосуды в проводящих пучках. Слабо развита и
корневая система, а корневые волоски отсутствуют. Почти все водные растения -
многолетники, размножающиеся вегетативно. Некоторые водные растения (наяда,
роголистник) опыляются под водой; у других цветки поднимаются над водой, где и
происходит опыление. Некоторые водные растения приспособились к периодическому
высыханию водоёмов (например, частуха, стрелолист, жеруха).
Во флоре СССР насчитывается св. 260 видов цветковых водных растений,
преимущественно однодольных. Семена и плоды распространяются птицами либо
водными течениями. Среди водных растений есть полезные; съедобны семена
водяного ореха, корневища сусака, зерновки злака манника и др. Семена и плоды
многих водных растений служат кормом для некоторых птиц; а отмершими остатками
водных растений часто питаются беспозвоночные животные, служащие пищей рыбам.
Водные растения играют роль в самоочищении бассейнов, хотя иногда (например,
элодея, некоторые виды рдестов) и сами могут быть вредными при сильном
разрастании их в водоёмах и особенно в водохранилищах. Для предупреждения
быстрого и нежелательного распространения заросли водных растений выкашивают
специальными водными косилками; скошенные водные растения иногда употребляют на
корм скоту. Многие водные растения разводят в аквариумах. К водным растениям
относятся также многие водоросли (например, зелёные и синезелёные), которые,
сильно разрастаясь, могут вызывать замор рыбы и зарастание каналов и прудов-
охладителей тепловых электростанций. Иногда для очищения каналов и других
водоёмов разводят растительноядные виды рыб (белый амур и белый толстолобик).
Для уничтожения водных растений используют также гербициды. Для водных растений,
служащих кормом для рыб, разработана специальная агротехника.
Лит.: Жизнь пресных вод СССР, под ред. В. И. Жадина, т. 2, М. - Л., 1949;
Скадовский С. Н., Экологическая физиология водных организмов, М., 1955;
Шмитхюзен И., Общая география растительности, пер. с нем., М., 1966; Новые
исследования по экологии и разведению растительноядных рыб. [Сб. ст.], М., 1968.
Водоросли
Ю.Е.Петров
Водоросли (Algae), группа низших, автотрофных, обычно водных, растений;
содержат хлорофилл и другие пигменты и вырабатывают органические вещества в
процессе фотосинтеза. Цветков и семян нет. Споры, как правило, лишены твёрдой
оболочки. Тело водорослей (слоевище, или таллом) по своему строению проще, чем у
мхов, папоротников и других наземных растений; часто отсутствует дифференциация
клеток на ткани; у самых примитивных водорослей (синезелёных) клетки лишены
оформленных ядер и хроматофоров; у части водорослей в клетках содержится помногу
ядер; есть водоросли неклеточного строения (ботридиум, сифоновые). Хроматофоры
у водорослей бывают пластинчатые, звездчатые, лентовидные, сетчатые, мелкие
дисковидные (последние характерны для высших представителей ряда типов
водорослей). У многих водорослей имеются плотные образования - пиреноиды и
пиреноидообразные тельца; у высших водорослей (почти всех бурых и большинства
красных) они отсутствуют. Клеточные оболочки состоят из целлюлозы, пектиновых
веществ, кремнийорганических соединений (диатомовые), альгина и фуцина (бурые).
Запасные вещества: крахмал, гликоген, полисахариды, реже масло. Насчитывается
около 30 тыс. видов водорослей. На основании различий в наборе пигментов,
особенностей морфологии и биохимии (состав клеточных оболочек, запасные
вещества) различают 10 типов (отделов) водорослей: синезелёные (Cyanophyta),
золотистые (Chrysophyta), пиррофитовые (Pyrrophyta), диатомовые
(Bacillariophyta), разножгутиковые, или жёлтозелёные (Xanthophyta), эвгленовые
(Euglenophyta), зелёные (Chlorophyta), харовые (Charophyta), бурые (Phaeophyta),
красные (Rhodophyta). Все типы водорослей эволюционировали в основном
независимо. Водоросли (по-видимому, зелёные) дали начало наземным растениям.
Размеры водорослей колеблются от долей микрона (кокколитофориды и некоторые
диатомовые) до 40 м (макроцистис). Многие водоросли одноклеточные; среди них
есть подвижные, совершающие скользящие движения (диатомовые, десмидиевые,
синезелёные), механизм передвижения которых окончательно не выяснен, и
водоросли, снабженные жгутиками, во многом подобные простейшим - жгутиковым, но
отличающиеся от них наличием хлорофилла и хроматофоров. Они могут утрачивать
хлорофилл (в темноте), становиться бесцветными и существовать за счёт
поглощения органических веществ, растворённых в воде; есть также виды
одноклеточных водорослей, способные, подобно простейшим, захватывать
органические частицы (некоторые пиррофитовые). Одноклеточные водоросли часто
при помощи слизи или выростов объединяются в колонии. Среди многоклеточных
водорослей наряду с крупными есть микроскопические; наиболее просто
организованные из них имеют вид разветвленных нитей, состоящих из одного ряда
клеток; другие имеют слоевища: корковидные, шнуровидные, шаровидные,
пластинчатые или кустистые с "листьями", снабженными жилками (саргассум). У
части синезелёных, зелёных и красных водорослей в слоевище откладываются
соединения кальция, и оно становится твёрдым. Водоросли лишены корней и
поглощают нужные им вещества из воды всей поверхностью. Крупные донные
водоросли имеют органы прикрепления - подошву (уплощённое расширение в
основании) или ризоиды (разветвленные выросты). У некоторых водорослей побеги
стелются по дну и дают новые слоевища.
Размножение водорослей - вегетативное, бесполое и половое. Многие одноклеточные
водоросли размножаются делением на две части. Крупные водоросли размножаются
вегетативно - частями слоевища или при помощи специальных почек (сфацеляриевые).
Некоторые многоклеточные водоросли не имеют полового размножения, у большинства
же образуются споры и гаметы либо в обычных клетках (зелёные водоросли, часть
красных), либо в особых образованиях - спорангиях и гаметангиях (бурые
водоросли); споры и гаметы бывают неподвижными (красные, конъюгаты) или
подвижными - со жгутиками. У водорослей наблюдаются все формы полового процесса:
изогамия, гетерогамия, оогамия и конъюгация (слияние протопластов двух
вегетативных клеток). Образующаяся в результате полового процесса зигота
делится сразу или после периода покоя. Одновременно в ней может происходить
мейоз. У примитивных водорослей одна и та же особь даёт гаметы или споры в
зависимости от внешних условий. У других водорослей функции бесполого и полового
размножения выполняют разные особи (спорофиты и гаметофиты); они могут
произрастать одновременно в одинаковых условиях (фурцелярия); одновременно, но
в разных местообитаниях (бангиевые); в одних и тех же местообитаниях, но в
разные сезоны. У ряда водорослей происходит строгое чередование гаметофита и
спорофита, которое принято называть "чередованием поколений". При этом у высших
водорослей зигота или прорастает на гаметофите, на нём же вырастает и спорофит
(ламинариевые), а гаметофит отмирает, или спора, не отделяясь от спорофита,
прорастает в гаметофит, который развивается на спорофите (фукусовые). Советский
специалист по водорослям - альголог М.М.Голлербах предложил для этого явления
термин "смена форм развития", наиболее верно отражающий существо процесса.
Спорофит у водорослей часто диплоиден (ядра содержат двойной набор хромосом), а
гаметофит гаплоиден (ядра с одинарным набором хромосом). В ряде случаев
гаметофит и спорофит находятся в одной ядерной фазе - оба гаплоидны (бангиевые)
или оба диплоидны (некоторые кладофоры, фукусовые).
Мелкие свободноплавающие водоросли входят в состав планктона и, развиваясь в
больших количествах, вызывают "цветение" (окрашивание) воды. Бентосные
водоросли прикрепляются ко дну водоёма или к другим водорослям. Есть водоросли,
внедряющиеся в раковины и известняк (сверлящие); встречаются (среди красных) и
паразитические. Крупные морские водоросли, главным образом бурые, образуют
нередко целые подводные леса. Большинство водорослей обитает от поверхности воды
до глубины 20-40 м, единичные виды (из красных и бурых) при хорошей
прозрачности воды опускаются до 200 м. Водоросли нередко в большом количестве
живут на поверхности и в верхних слоях почвы, одни из них усваивают атмосферный
азот, другие приспособились к жизни на коре деревьев, заборах, стенах домов,
скалах. Микроскопические водоросли вызывают красное или жёлтое "окрашивание"
снега высоко в горах и в полярных районах. Некоторые водоросли вступают в
симбиотические отношения с грибами (лишайники) и животными.
Водоросли - главные производители органических вещества в водной среде. Около
80% всех органических веществ, ежегодно создающихся на земле, приходится на
долю водорослей и других водных растений. Водоросли прямо или косвенно служат
источником пищи для всех водных животных. Известны горные породы (диатомиты,
горючие сланцы, часть известняков), возникшие в результате жизнедеятельности
водорослей в прошлые геологические эпохи. Водоросли участвуют в образовании
лечебных грязей. Некоторые, в основном морские, употребляются в пищу (морская
капуста, порфира, ульва). В приморских районах водоросли идут на корм скоту и
удобрение. В ряде стран водоросли культивируют для получения большого
количества биомассы, идущей на корм скоту и используемой в пищевой
промышленности. Многие водоросли - важный компонент процесса биологической
очистки сточных вод. Из водорослей получают: студне и слизеобразующие вещества -
агар-агар (анфельция, гелидиум), агароиды (филлофора, грацилярия), карраген
(хондрус, гигартина, фурцелярия), альгинаты (ламинариевые и фукусовые),
кормовую муку, содержащую микроэлементы и иод. Водоросли широко применяют в
экспериментальных исследованиях для решения проблем фотосинтеза и выяснения
роли ядра и других компонентов клетки. Предпринимаются попытки использовать
некоторые быстро размножающиеся и неприхотливые водоросли (например, хлореллу,
которая быстро и в большом количестве синтезирует белки, жиры, углеводы,
витамины и достаточно полно поглощает вещества, выделяемые человеком и
животными) для создания круговорота веществ в обитаемых отсеках космических
кораблей. Наука о водорослях называется альгологией.
Лит.: Арнольди Водоросли М., Введение в изучение низших организмов, М. - Л.,
1925; Курсанов Л. И. и Комарницкий Н. А., Курс низших растений, 3 изд., М.,
1945; Воронихин Н. Н. и Шляпина Е. Водоросли, Водоросли, в кн.: Жизнь пресных
вод СССР, под ред. Водоросли И. Жадина, т. 2, М. - Л., 1949, гл. 19;
Определитель пресноводных водорослей СССР, в. 1 - Голлербах М. М. и Полянский
Водоросли И., Пресноводные водоросли и их изучение, М., 1951; Чэпмен Водоросли,
Морские водоросли и их использование, пер. с англ., М., 1953; Определитель
низших растений, под ред. Л. И. Курсанова, т. 1-2, Водоросли, М., 1953; Зинова
А. Д., Определитель зеленых, бурых и красных водорослей южных морей СССР, М. -
Л., 1967; Oltmanns F., Morphologie und Biologie der Algen, 2 Aufl., Bd 1-3,
Jena, 1922- 23: Fritsch F. Е., The structure and reproduction of the algae, v.
1-2, Camb., 1935-45; Smith G. М., Manual of phycology, L., 1951; Fott B.,
Algenkunde, Jena, 1959; Chapman V. J., The algae, L., 1962.